WELCOME TO OPEFE ARCHIVES
WELCOME TO OPEFE ARCHIVESSerrasalmidae
Pirambeba Dorada or Piranha Negra
Serrasalmus spilopleura
KNER 1858
This species was originally described from Rio Guapore, Bobota, Mato Grosso, Brazil (Kner 1858:166 [6] [Sitzungsber. Akad. Wiss. Wien v. 32 (no. 22)). However, Jegu 2001 places populations of S. spilopleura as widespread according to the distribution below. Because an error in misidentification was made between S. spilopleura and S. eigenmanni in Jegu and colleague in 2001, spilopleura is found only in Bolivia according to Hubert et al. 2007, 2010. On the above left corner is the only photo I have of a correctly identified specimen (per M. Jegu) of S. spilopleura. Below is the living example (same as pictured above) in an aquarium. Video by Martin Reilly;
The maps shown below are approximate range of certain species. It does not show the complete range. No other species has given me so much reason to pause and consider as this one and S. maculatus. The problem centers on the description and rehabilitation of the species.
This rehabilitation places the true S. spilopleura from the Guaporé Basin of Western Brazil (Bolivian side rio Itenez) only (see). The Central Amazon fishes are S. maculatus. To further add to this confusion, the fish from the Pantanal region have not been rehabilitated but is being recognized as S. maculatus as well. So hobbyists are cautioned about using either name on the species from that region until more is learned.
REFERENCES FROM CAS
Serrasalmus spilopleura Kner [R.] 1858:166 [6] [Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften. Mathematisch-Naturwissenschaftliche Classe v. 32 (no. 22); Rio Guaporé, Bobota, Mato Grosso, Brazil. Lectotype: NMW 57085. Paralectotypes: NMW 16344 (1, dry), 79457-59 (1, 1, 1, all dry). Appeared in more detail in Kner 1860:43 [p. 35 of separate], Pl. 5 (fig. 11). Lectotype designated by Jégu & Santos 2001:125. •Valid as Serrasalmus spilopleura Kner 1858 -- (Ortega & Vari 1986:9, Cestari & Galetti 1992, Gómez & Chebez 1996:51, Britski et al. 1999:66, Butí & Cancino 1999:71, Nakayama et al. 2000:149, Jégu & Santos 2001:119, Lasso et al. 2001:96, Jégu in Reis et al. 2003:192, López et al. 2003:32, Casciotta et al. 2003:116, Menni 2004:78). Current status: Valid as Serrasalmus spilopleura Kner 1858.
|
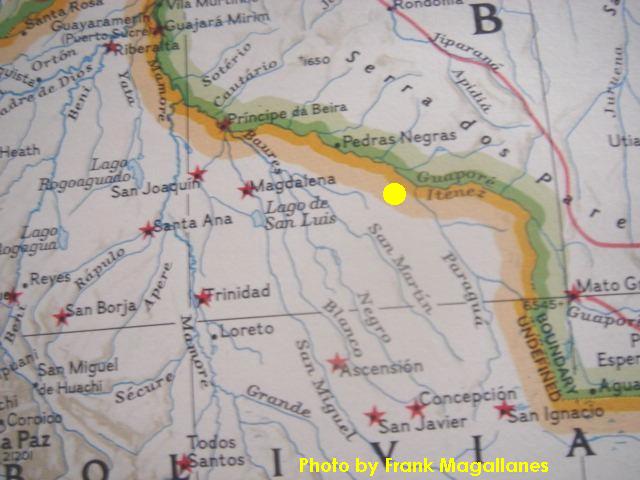
Map showing rio Itenez where S. spilopleura is found. It begins at the mouth of the Mamore. |
MAXIMUM SIZE - According to Fishbase.org (expect errors).
20.0 cm SL.
DISTRIBUTION
Guaporé River basin, Paraná River basin: Argentina and Brazil.
REFERENCES
Jégu, M. and G. M. dos Santos 2001 Mise au point ŕ propos de Serrasalmus spilopleura Kner, 1858 et réhabilitation de S. maculatus Kner, 1858 (Characidae: Serrasalminae). Cybium v. 25 (no. 2): 119-143. (1) Antenne IRD, Laboratoire d'Ichtyologie générale et appliquée, M.N.H.N., 43 rue Cuvier, 75231 Paris Cedex 05 France. (2) Laboratorio de Ictiologia, I.N.P.A., C.P.B.A., Cx P. 478, 69083-970 Manaus, A.M. Brézil.
Hubert N. & Renno J. F. 2010 Description of a new Serrasalmus species, Serrasalmus odyssei n. sp. [pp. 52-59]. In: Evolution of the neotropical ichthyofauna -- molecular and evolutionary perspectives about the origin of the fish communities in the Amazon. VDM Publishing House, Verlag Dr. Muller, Saarbucken, Germany. Evolution of the Neotropical Ichthyofauna.: 95 pp.
Aylton Saturnino Teixeira; Celeste Mutuko Nakayama; Jorge Ivan Rebelo Porto; Eliana Feldberg, (2006) Esterase-D and chromosome patterns in Central Amazon piranha (Serrasalmus rhombeus Linnaeus, 1766) from Lake Catalăo, Genet. Mol. Biol. v.29 n.3 Săo Paulo.
Allendorf FW and Utter FM (1979) Population genetics. In: Hoar WS, Randall DJ and Brett JR (eds) Fish Physiology, v. 8. Academic Press, New York, pp 407-454.
Fink, W. L. 1993 (18 Aug.) Revision of the piranha genus Pygocentrus (Teleostei, Characiformes). Copeia 1993 (no. 3): 665-687.
Alonso SK and Alfenas AC (1998) Isoenzimas na taxonomia e na genética de fitonematóides. In: Alfenas AC (ed) Eletroforese de Isoenzimas e Proteínas Afins. Editora da Universidade Federal de Viçosa, Viçosa, pp 525-539.
Anti AB (2000) Caracterizaçăo de germoplasma de soja e de feijăo através de eletroforese de isoenzimas da semente. Bragantia 59:139-142
Dissertaçăo de Mestrado aprovada pela PUCRS em 2001 com o título: Aspectos da biologia da piranha Serrasalmus spilopleura (Characidae) no Município de Uruguaiana, oeste do Estado do Rio Grande do Sul, Brasil. Rodrigo C. and P. Beheregaray Porto Alegre - RS - Brasil.
Froese,
R. and D. Pauly. Editors. 2002. FishBase.
World Wide Web electronic publication. 13 November 2002.
Cavicchioli, M; Nakatani, K; Shibatta, OA (1997): Morphometric variation of larvae and juveniles of the piranhas Serrasalmus spilopleura and S. marginatus (Characidae: Serrasalminae) of the Paraná basin, Brazil. Ichthyol. Explor. Freshwaters 8 (2), 97-106.
Fink, W.L. 1993 Revision of the piranha genus «Pygocentrus» (Teleostei, Characiformes). Copeia 1993(3):665-687
Spix, J.B. von & Agassiz, L. 1829 Selecta genera et species piscium quos in itinere per Brasiliam annos MDCCCXVII-MDCCCXX jussu et auspiciis Maximiliani Josephi I... collegit et pingendos curavit Dr. J.B. de Spix... Monachii. This is a book. Part 1:i-xvi+i-ii+1-82, pl.1-49; Part 2:83-138, pl.49-101.
CENTOFANTE, L.; PORT, J.I.R. ; FELDBERG, And 2002. Chromosomal polymorphism in Serrasalmus spilopleura Kner, 1858 (Characidae, Serrasalminae) from Central Amazon Basin. Caryologia , 55(1):41-49.
PORT, J. I. R. ; NAKAYAMA, C. M.; FELDBERG, And 2002. Comparative cytogenetic study of five piranha species (Serrasalmus, Serrasalminae) from the Amazon Basin. Genetica.
NAKAYAMA, C.M.; JEGU, M.; FELDBERG, And & PORT, J.I.R. (1993). Description of two sympatric and syntopic cytotypes of piranhas of Serrasalmus rhombeus - complex (Teleostei, Serrasalmidae) from the Central Amazon basin (Amazon, Brazil).
CENTOFANTE, L.; PORTO, J.I.R. & FELDBERG, E. (1999) Análise cromossômica na espécie de piranha Serrasalmus spilopleura (Characiformes, Serrasalmidae) do lago Catalăo, Amazonas. Resumos do I Simpósio de Pós-Graduaçăo do INPA/FUA: 14.
Sazima, I. and S. Guimaraes. 1987. Scavenging on human corpses as a source for stories about man-eating piranhas. En. Biol. Fishes 20: 75-77.
Casciotta, J., A. Almirón and J. Bechara 2003 Los peces de la laguna Iberiá. Colección Universitaria Ciencias Naturales. Peces Iberá: 1-203
Menni, R. C. 2004 (5 Mar.) Peces y ambientes en la Argentina continental. Monogr. Mus. Argentina Cienc. Nat. No. 5: 1-316.
Reis, R. E., S. O. Kullander and C. J. Ferraris, Jr. 2003 Check list of the freshwater fishes of South and Central America. Check list Freshw. Fishes South & Cent. Amer. 2003: i-xi + 1-729.
USE YOUR BACKSPACE TO RETURN OR CLICK HERE TO RETURN SERRASALMUS LIST
The OPEFE web site and its contents; is disclaimed for purposes of Zoological Nomenclature in accordance with the International Code of Zoological Nomenclature, Fourth Edition, Article 8.3 and 8.4. No new names or nomenclature changes are available from statements at this web site.
Copyright© 1994-2012 Oregon Piranha Exotic Fish Exhibit (The OPEFE fish exhibit is permanently CLOSED as of 2000) Sutherlin, Oregon. Information posted on this web site is archival data on fish scientific classifications and other information. DISCLAIMER: The copyrighted material may not be used for any purpose other than private study, scholarship or research. Cited information requires credit and this link www.opefe.com. All rights reserved. All images shown (unless otherwise noted) is property of OPEFE.
UPDATED: 11/14/2014